
Druck-Version
Hintergrund
T-urf13 revisited: Ein 'intelligent designtes' Protein ohne Designer
Über die zufällige Entstehung eines nicht reduzierbar komplexen Systems
Ein beliebtes Argument von Evolutionsgegnern lautet, biologische Systeme seien derart komplex, dass sie nicht in hinreichend kleinen Zwischenschritten evolvieren könnten. Es sei erst recht sehr unwahrscheinlich, dass spezifisch komplexe Strukturen und Funktionen auf einen Schlag entstünden. Dies, so die Behauptung, könne man bereits anhand einzelner Proteine zeigen, die hochgradig spezifisch für bestimmte Substrate, Wechselwirkungen und Molekularprozesse "ausgelegt" seien und einen sehr individuellen strukturellen Aufbau zeigten.[1]
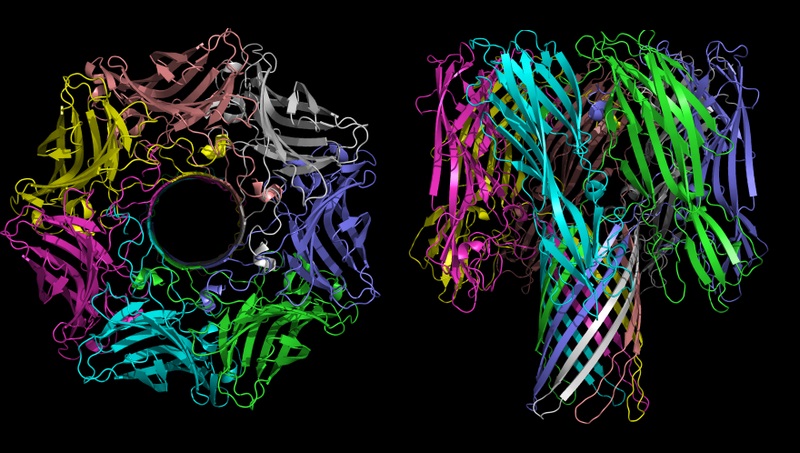
Titelbild: α-Hämolysin aus der Gruppe porenbildender Toxine. Bildquelle: Bassophile, Representation of Alpha Haemloysin from Staph aureus, created from PDB file 7ahl, using PyMol, CC BY-SA 3.0.
{kind=link}
Das Stichwort dazu lautet nicht-reduzierbare Komplexität. Ein Beispiel ist der im Titelbild dargestellte, porenbildende Protein-Komplex namens α-Hämolysin, welcher aus sieben Protein-Untereinheiten besteht. Dieses Toxin des Bakteriums Staphylococcus aureus bewirkt eine Hämolyse der roten Blutkörperchen (Zellzerstörung). Die sieben Untereinheiten tragen mit jeweils zwei β-Faltblatt-Strukturen, die im Kreis angeordnet sind, und hydrophilen Aminosäuren im Inneren zur Ausbildung einer röhrenförmigen Pore (β-Fass-Struktur) bei.
Entgegen der Behauptung der Evolutionsgegner zeigen wir, dass derlei Strukturen (hier: ein komplettes Protein mit mehreren spezifischen Bindestellen und Strukturelementen, die mit weiteren Einheiten einen komplexen, funktionellen Ionenkanal bilden) bereits in kurzer Zeit de novo evolvieren können. Ein besonders eindrückliches und schönes Beispiel steckt hinter der Geschichte des Proteins T-URF13, das wir hier vorstellen.
Die Vorgeschichte: Heterosis-Mais
Anfang des 20. Jahrhunderts versuchte man in den USA, robuste Mais-Sorten zu züchten, die auch unter schwierigen Bedingungen gut wachsen. Bald stellte sich heraus, dass insbesondere Heterosis-Pflanzen - mischerbige Pflanzen mit genetisch deutlich unterschiedlichen Eltern - am widerstandsfähigsten sind. Das Problem bei der Anzucht ist jedoch, dass der Heterosis-Grad, also das erwünschte, hohe Ausmaß an Mischerbigkeit, mit jeder nachfolgenden Generation sinkt, sodass der physiologische Vorteil ziemlich bald verschwindet.
Um diesem Effekt zu begegnen, züchtete man in einer der beiden vorgesehenen elterlichen Linien eine Pollensterilität. Daher sind diese Pflanzen auf Fremdbestäubung angewiesen. Somit konnte man Jahr für Jahr wieder "frischen" Heterosis-Saatmais erzeugen. Dies gelang mit verschiedenen Zuchtlinien.
Pollensteriler Mais
Viele der pollensterilen Linien wurden genetisch untersucht, um herauszufinden, worauf die Pollensterilität beruht. Es war zu erwarten, dass irgendwo im Genom ein Gen ausgefallen war, das für die Pollenreifung essenziell ist. Überraschenderweise erwies sich beim Texas Male-Sterile Maize (CMS-T) ein gänzlich anderer Effekt als verantwortlich für die Pollensterilität: Im Genom der Mitochondrien wurde ein komplett neues Gen gefunden, welches für die Sterilität verantwortlich war. Seine Entdecker nannten das Gen T-urf13. Das entsprechende Protein (T-URF13, manchmal auch URF13) wird nicht kursiv, dafür aber groß geschrieben.
Struktur und Funktion des T-URF13-Proteins
Umfangreiche Folgearbeiten [2]–[14] entschlüsselten Herkunft, Struktur und Wirkungsweise von T-URF13. Es handelt sich um ein Porenprotein in der inneren Wand der Zellkraftwerke (innere Mitochondrienmembran). Dort liegt es als Verbund (Multimer) vor – meist aus vier gleichen Bausteinen (Monomeren). Jedes Monomer besitzt drei die Membran durchdringende, spiralförmige Abschnitte (α-Helices), die sich gemeinsam zu einer chemisch regulierbaren Schleuse für Elektrolyte (einem gesteuerten Ionenkanal) zusammenlagern.
Zu einem ganz bestimmten Zeitpunkt der Blütenreifung wird dieser Kanal aktiv: Er bewirkt gezielt das Absterben derjenigen Zellen, die für die Entwicklung der Pollen zuständig sind (Tapetumzellen). Da die restliche Pflanze von diesem Vorgang unberührt bleibt, wächst sie vollkommen intakt und vital weiter – sie ist danach lediglich pollensteril.
T-URF13 vereint in seiner modularen Struktur vier funktionelle Kernmerkmale, deren molekulares Zusammenspiel die Gesamtfunktion des Proteins überhaupt erst ermöglicht:
- Komplexbildung: Die T-URF13-Monomere besitzen spezifische Kontaktbereiche, über die sie sich in der Zellmembran zu einem multimeren Proteinkomplex zusammenlagern.
- Kanalsteuerung: Dieser Proteinkomplex bildet eine Pore (einen Ionenkanal) und wechselt zwischen zwei definierten Strukturen (offen und geschlossen). Dieses Öffnen und Schließen (Gating) setzt eine Reihe spezifisch-struktureller Anforderungen voraus.
- Signalsteuerung: Der Komplex besitzt eine spezifische Bindungsstelle für Signalmoleküle (Liganden), die das Öffnen oder Schließen des Kanals auslösen.
- Genexpression: Das Protein muss in ausreichend hoher Konzentration in der Membran vorliegen, da erst eine kritische Dichte aktivierter Kanäle die Pollensterilität einleitet.
T-URF13 erfüllt somit par excellence die Definition eines "nichtreduzierbar komplexen" Systems nach Michael Behe, weil die biologische Funktion das gleichzeitige Vorhandensein mehrerer Teile (bzw. der dafür verantwortlichen strukturellen Voraussetzungen) erfordert.
Die molekulare Herkunft von T-URF13
Nach der Identifikation und Sequenzierung des kodierenden Bereichs lag die Vermutung nahe, T-URF13 stamme von einem bereits existierenden Ionenkanal ab, dessen Aktivität sich durch Mutationen verändert hatte. Eine Ähnlichkeitssuche in den Sequenzdatenbanken (BLAST) widerlegte dies jedoch (Abb. 2 und 3): Das T-urf13-Gen ist eine molekulare Chimäre aus verschiedenen Abschnitten. Es setzt sich zusammen aus:
- einer ehemals nicht abgelesenen (intergenischen) Region des Mitochondrien-Erbguts (Abb. 2, gelb),
- einem Teil des mitochondrialen Gens für die 26S-rRNA (Abb. 2, grün), das im gesunden Organismus zwar abgelesen, aber niemals (und zwar in keinem Lebewesen) in ein Protein translatiert wird.
- der Bruchstelle (Abb. 2, rot), die bei der unpräzisen Fusion des gelben und des grünen DNA-Fragments als neues Verbindungsstück entstand.
Ferner ist t-urf13 in die 3547 bp lange Region TURF 2H3 eingebettet. Diese entstand durch Rekombinationen von Abschnitten des mitochondrialen 26S-rRNA-Gens, des atp6-Gens und eines Chloroplasten-tRNA-Arg-Gens [2]. Dem t-urf13-Leseraster wurde dabei ein funktioneller atp6-Promotor vorgeschaltet, der für eine konstitutiv hohe Transkriptionsrate sorgt.

Abb. 2 Proteinkodierender Bereich des T-urf13-Gens. Dargestellt ist der offene Leserahmen vom Start- (ATG) bis zum Stop-Codon (TGA). Die Farbkodierung markiert die genomische Herkunft der DNA-Segmente: Der gelbe Abschnitt (Nukleotid 1–263) umfasst die ehemals nicht transkribierte, intergenische Region. Der rote Abschnitt (Nukleotid 264–285) kennzeichnet die direkt bei der unpräzisen Fusion entstandene Bruchstelle. Der grüne Abschnitt (Nukleotid 286–348) markiert das Fragment, das aus der ehemals rein strukturellen Region des mitochondrialen 26S-rRNA-Gens stammt.

Abb. 3 Primärstruktur des T-URF13-Proteins (IUPAC-Einbuchstabencode). Hier ist die fertige Aminosäurekette zu sehen. Sie besteht aus 115 Bausteinen. Die Farben zeigen, woher die Teile ursprünglich stammen: Der gelbe Anfang sorgt dafür, dass sich das Protein in der inneren Mitochondrienmembran (den Kraftwerken der Zelle) verankert. Der rote Teil ist das neu entstandene "Verbindungsstück" (EGGGRSN), das offenbar im Zuge der Kombination der beiden flankierenden Teile (gelb und grün) entstand. Der grüne Teil stammt aus dem Bereich, der vorher nie für Proteine genutzt wurde.
Wichtig: Kein einzelner dieser Abschnitte besitzt für sich genommen den Bauplan für eine Pore. Erst wenn sich mehrere der fertigen Proteinketten kreisförmig in der Membran zusammenlagern – ähnlich den Dauben eines Weinfasses –, entsteht die Porenfunktion. Sie ist eine emergente Eigenschaft – ein völlig neues Merkmal, das erst durch das Zusammenspiel der zuvor zweckfremden Fragmente entsteht.
Das Faszinierende an diesem Entstehungsprozess ist, dass diese Ursprungssequenzen in der gesamten evolutionären Geschichte der Eukaryoten niemals proteinkodierend waren – mit Ausnahme des kurzen atp6-Fragments, das zuvor jedoch für ein funktionell völlig anderes Protein kodierte und nichts mit einem Porenprotein zu tun hat.
Handelte es sich unter diesen Voraussetzungen nicht um ein astronomisch unwahrscheinliches Ergebnis? Nicht nach den Prinzipien der Proteinfaltung: Die strukturellen Anforderungen an stabile Helices sind überraschend gering. Arbeiten an künstlichen Proteinen zeigen, dass bereits ein passendes, binäres Muster aus hydrophoben und hydrophilen Aminosäuren genügt, um stabile Sekundärstrukturen entstehen zu lassen (vgl. Hecht 1990; Kamtekar et al. 1993). Bei T-URF13 führte dieses zufällige Muster dazu, dass sich die Fragmente in der Membran verankerten und spontan zu einem oligomeren Porenkomplex zusammenlagerten.
Der Entstehungsprozess
Der evolutionäre Ursprung des Gens T-urf13 ist exzellent dokumentiert, da es exklusiv in der Maissorte CMS-T vorkommt und die Ursprungssequenzen in den unveränderten Ausgangslinien exakt lokalisiert werden konnten. Damit liegen lückenlose empirische Belege für die natürliche Entstehung des Systems vor.
Diese Entstehung war kein plötzlicher, singulärer "Sprung". Stattdessen vollzog sich der Prozess schrittweise über mehrere aufeinanderfolgende Rekombinationsereignisse, die im mitochondrialen Genom abliefen. Bei T-urf13 basierte diese Neuordnung auf mitochondrialen Doppelstrangbrüchen: Bei deren Reparatur (über fehleranfällige Mechanismen wie non-homologous end joining oder single strand annealing) wurden die oben beschriebenen DNA-Fragmente zufällig fusioniert. Da sich die Sequenzen an den Verknüpfungsstellen oft regellos verändern, entstand zudem ein Sequenzabschnitt (der "rote Bereich", Abb. 2 und 3), für den sich kein evolutionärer Vorläufer finden lässt.
Da all diese vorbereitenden Umlagerungen mangels eines aktiven Promotors und eines durchgehenden Leserahmens phänotypisch vollkommen "stumm" blieben, waren sie für die natürliche Selektion unsichtbar und konnten nicht weggefiltert werden. Erst der finale Rekombinationsschritt – der Anschluss an den funktionellen atp6-Promotor – enthüllte dieses im Genom unsichtbar angewachsene Potenzial schlagartig und überführte die ehemals funktionslosen Fragmente in ein neues, komplexes Proteinsystem.
Trotter et al. (2014) zeigen in ihrem populationsgenetischen Modell, dass solche phänotypisch stummen (oder kryptischen) Variationen im Hintergrund einen gigantischen kombinatorischen Sequenzraum bereitstellen. Sie ermöglichen es aufgrund der schieren Menge an potenziellen Kombinationen, breite Fitnesstäler zu durchqueren und nichtreduzierbar komplexe Systeme hervorzubringen. Die Entstehung von T-urf13 untermauert dies eindrucksvoll.
Ein starkes Argument gegen Intelligent Design
Die Entstehung dieses Systems widerlegt gängige Argumente gegen die Evolutionstheorie. Nach Ansicht des Intelligent-Design-Vertreters Michael Behe (2007) ist es praktisch unmöglich, dass Proteine mit mehr als zwei spezifischen Bindestellen auf natürlichem Weg evolvieren. Selbst eine Doppelmutation sei im Laufe von Jahrmilliarden nur ein einziges Mal unter Abertrillionen von Einzellern möglich – erst recht nicht bei komplexen Kulturpflanzen wie dem Mais. Behes Berechnungen (am Beispiel der Chloroquin-Resistenz des Malaria-Erregers Plasmodium falciparum) erweisen sich jedoch in ihren Voraussetzungen als fehlerhaft.
Das T-URF13-Protein liefert den direkten Gegenbeleg: Ein System, das nach Behes Definition als "nicht reduzierbar komplex" gilt, entstand hier innerhalb kürzester Zeit über ganz reale, nachweisbare Evolutionsmechanismen und sprengt die von ihm postulierten Grenzen der Naturwissenschaft fundamental.
Warum der Vergleich mit technischem Design hinkt
Dieser Vorgang lässt sich mit einem technischen Gleichnis verdeutlichen: Stellen Sie sich vor, Bilddateien auf einem PC werden durch einen Kopierfehler beschädigt. Die Fragmente kombinieren sich neu, etliche Bits verändern sich regellos, und es entsteht eine kleinere Datei. Diese ist nun keine Bilddatei mehr, sondern ein funktionierendes Programm, das plötzlich Musikdateien abspielen kann.
In der Softwareentwicklung ist so etwas unmöglich. In der biologischen Evolution hingegen zeigt das Beispiel T-urf13, dass technische und biologische Systeme nur sehr begrenzt vergleichbar sind. Die menschliche Intuition ist bei der Betrachtung der Natur allzu oft kein guter Ratgeber.
Mögliche Gegenargumente
Aus der ID-Perspektive kann man mit nachstehenden Standard-Einwänden rechnen, die noch kurz abgehandelt seien. Eine ausführliche Erwiderung auf die Gegenargumente des evangelikalen Discovery-Instituts findet sich an dieser Stelle (BEYER et al. 2022).
1. Homologie bzw. Sequenzähnlichkeiten liefern keinen Entstehungsmechanismus. Niemand hat valide bewiesen, dass T-urf13 durch rekombinante Evolutionsereignisse entstanden ist.
Dieser Einwand verkennt die methodologischen Grundlagen der Naturwissenschaften: Empirische Disziplinen arbeiten nicht mit mathematischen Beweisen, sondern mit stützenden oder falsifizierenden Indizienketten (Belegen). Zudem entlarvt der Einwand einen methodischen Widerspruch: Evolutionsexperimente, in denen komplexe Neuerungen entstehen, sind genau das, was ID-Vertreter seit jeher fordern. Die dokumentierte Zuchtgeschichte des CMS-T-Maises liefert aus folgenden Gründen diesen empirischen Entstehungsnachweis:
- Das Gen fehlt bei allen wilden Verwandten und ursprünglichen Maissorten.
- Es verursacht spezifisch die Pollensterilität und trat exakt in den darauf selektierten Zuchtlinien auf.
- Es lässt sich sequenzgenau auf bekannte mitochondriale DNA-Quellen (das 26S-rRNA-Gen und Genflanken) zurückführen; die molekularen Bruchstellen der Rekombination sind exakt nachgewiesen.
Die für die Entstehung der Neuerungen bzw. der Rearrangements erforderlichen zellulären Mechanismen sind seit Jahrzehnten bekannt und gut verstanden. Die De-novo-Entstehung aus vorhandenen genomischen Ressourcen ist somit die einzig plausible Erklärung.
2. Das Auftreten von T-urf13 in Zuchtlinien beweist kein ungerichtetes Evolutionsereignis. Ein Züchtungsexperiment wird von Menschen kontrolliert – der Züchter fungierte hier als "Intelligent Designer".
Diese Interpretation verdreht die biologischen Fakten des Züchtungsprozesses. Der menschliche Züchter hat dieses Gen weder im Labor geplant, noch am Computer designt oder künstlich in das Genom eingefügt. Die molekulare Entstehung der Sequenz war ein unvorhergesehenes, rein stochastisches (zufälliges) Nebenprodukt natürlicher zellulärer Mechanismen.
Der Züchter war dabei nicht der "Schöpfer" der genetischen Information, sondern stellte lediglich den künstlichen Selektionsfaktor dar. Das Experiment zeigt exakt das Zusammenspiel der evolutionären Kernmechanismen: Mutationen (mitochondriale Rekombinationen) erzeugen eine neue Eigenschaft, die dann durch veränderte Umweltbedingungen (die Selektion durch den Züchter) im Genpool fixiert werden.
3. Die Herkunft von T-urf13 ist ungeklärt. Wahrscheinlich stammt es von einem anderen ("intelligent programmierten") Vorläufer ab, der nur minimal mutieren musste, damit das beobachtete Ergebnis herauskommt.
Wie dargelegt, entbehrt dieser Einwand angesichts moderner Datenbestände jeder Grundlage. Da globale Sequenzdatenbanken mittlerweile eine immense Vielfalt an Genomen von Wildtypen und Verwandten des Maises umfassen, lässt sich das Fehlen eines solchen Vorläufergens mit an Sicherheit grenzender Wahrscheinlichkeit annehmen. BLAST-Analysen zeigen keine signifikanten Treffer für die gesamte chimäre Sequenz außerhalb des spezifischen CMS-T-Mais-Mitochondrioms. Die Postulierung eines hypothetischen, funktionellen Vorläufergens bleibt damit eine reine Schutzbehauptung ohne wissenschaftliche Evidenz.
4. Die DNA-Elemente, aus denen T-urf13 zusammengesetzt ist, haben vom Designer / Schöpfer eine "versteckte Protein-Kodierungskapazität" mitbekommen.
Erstens handelt es sich bei einer solchen "versteckten Kodierungskapazität" um ein klassisches Ad-hoc-Argument (Immunisierungsstrategie): Sie wird nur im Nachhinein postuliert, ist empirisch nicht messbar und entzieht sich jeder Falsifikation.
Zweitens widersprechen die evolutionären Spuren in den Ursprungssequenzen dieser Hypothese. Vergleicht man die kodierenden Bereiche (open reading frames, ORFs) etablierter Proteingene über verschiedene Spezies hinweg, findet man fast ausschließlich InDels (= Einschübe oder Auslassungen) von 3, 6, 9 usw. Basen (Vielfache des Triplett-Codons), weil sie das Leseraster nicht zerstören. In nicht-kodierenden Regionen und rRNA-Genen hingegen findet man InDels aller Längen, da kein Selektionsdruck auf ein Protein-Leseraster existiert.
Hätten diese Regionen eine "vorausgeplante" Proteinkodierungskapazität besessen, hätte das Leseraster in den Vorläuferorganismen über Jahrmillionen durch konservierenden Selektionsdruck stabil gehalten werden müssen (da Verschiebungen des Leserasters – sogenannte Frameshifts – das resultierende Protein unbrauchbar gemacht hätten). Dass T-urf13 aus diesen unveränderten Sequenzen ohne die "3er-Regel" entstand, beweist, dass es sich um die De-novo-Entstehung einer Funktion aus einem reinen Zufalls-Leseraster handelt – ein stochastisches Nebenprodukt der mitochondrialen Rekombination, kein vorgefertigtes Design.
5. Die Funktion von T-URF13 ist destruktiv, nicht konstruktiv.
Hier wird der biologische Kontext mit der nichtreduzierbar komplexen Struktur verwechselt: Was zählt, ist die Tatsache, dass das T-URF13-Protein einen spezifischen Aufbau hat, spezifische Bindestellen für Liganden und eine spezifische molekulare Funktion.
6. Zur Entstehung des T-urf13 auf dem Mitochondrien-Genom waren mehrere Rekombinationsereignisse nötig, was das Szenario noch unwahrscheinlicher macht.
Wenn, wie bereits gesagt, das Gen nicht durch ein Wunder entstanden sein soll, lautet der Schluss auf die beste Erklärung nach wie vor, dass es durch eine Reihe von Gen-Rekombinationen entstand. Wie erläutert sind die dafür erforderlichen Prozesse bekannt und mechanismisch sehr gut verstanden. Oder wollen uns die ID-Vertreter Glauben machen, dass eine versteckte intelligente Macht Anfang des 20. Jahrhunderts den Züchtern "in die Karten spielte"? Das ist kein Schluss auf die beste Erklärung, das ist Obskurantismus. Im Übrigen ist das Argument zur Unwahrscheinlichkeit, wie Trotter et al. (2014) darlegen, nicht haltbar.
7. Der transkribierte Teil von T-urf13, der den gesteuerten Ionenkanal liefert, ist nur ein kleiner Teil des "de novo"-Gens. Außerdem ist ein Teil des mitochondrialen atp6 beteiligt, und das ist proteinkodierend.
Das atp6-Gen lieferte im Wesentlichen den Promotor sowie ein kleines, flankierendes Sequenzfragment. Ein Promotor ist eine reine regulatorische Andockstelle für Enzyme und wird ohnehin niemals in ein Protein translatiert. Das kurze, mitübertragene atp6-Codon-Fragment kodierte im Ursprungsprotein zudem für ein völlig anderes Signalpeptid und besaß keinerlei Porenfunktion oder Relevanz für das spätere Gating. Die funktionellen Schlüsselelemente des Ionenkanals stammen nachweislich aus den ehemals nicht-kodierenden Bereichen.
8. Es gibt eine lebhafte Debatte in den USA um T-urf13. ID-Vertreter haben gezeigt, dass die Argumentation nicht sticht.
Die wesentlichen Einwände haben wir oben und inzwischen auch auf der amerikanischen Internetplattform "Panda's Thumb" abgehandelt. Zudem wird die "kontroverse Diskussion" nur in Blogs und auf christlichen Internet-Seiten geführt, nicht in der Fachwelt. Die ID-Vertreter sind somit aufgefordert, ihre Daten und Argumente – sofern sie funktionierende besitzen – in begutachteten wissenschaftlichen Fachzeitschriften (peer-reviewed journals) zu publizieren.
Literatur
[1] Behe M (2007) The edge of evolution. Free Press, New York.
[2] Dewey RE, Levings CS 3rd, Timothy DH (1986) Novel recombinations in the maize mitochondrial genome produce a unique transcriptional unit in the Texas male-sterile cytoplasm. Cell 44(3): 439-449.
[3] Stamper SE, Dewey RE, Bland MM, Levings CS 3rd (1987) Characterization of the gene urf13-T and an unidentified reading frame, ORF 25, in maize and tobacco mitochondria. Curr Genet. 12(6): 457-63.
[4] Dewey RE, Timothy DH, Levings CS (1987) A mitochondrial protein associated with cytoplasmic male sterility in the T cytoplasm of maize. Proc Natl Acad Sci U S A. 84(15):5374-8.
[5] Braun CJ, Siedow JN, Williams ME, Levings CS 3rd (1989) Mutations in the maize mitochondrial T-urf13 gene eliminate sensitivity to a fungal pathotoxin. Proc Natl Acad Sci U S A. 86(12): 4435-9.
[6] Braun CJ, Siedow JN, Levings CS 3rd (1990) Fungal toxins bind to the URF13 protein in maize mitochondria and Escherichia coli. Plant Cell. 2(2):153-61.
[7] Levings CS 3rd. 1990 The Texas cytoplasm of maize: cytoplasmic male sterility and disease susceptibility. Science 250(4983):942-7.
[8] Korth KL, Kaspi CI, Siedow JN, Levings CS 3rd (1991) URF13, a maize mitochondrial pore-forming protein, is oligomeric and has a mixed orientation in Escherichia coli plasma membranes. Proc Natl Acad Sci U S A. 88(23):10865-9.
[9] Levings CS 3rd, Siedow JN (1992) Molecular basis of disease susceptibility in the Texas cytoplasm of maize. Plant Mol Biol. 19(1):135-47.
[10] Kaspi CI, Siedow JN (1993) Cross-linking of the cms-T maize mitochondrial pore-forming protein URF13 by N,N'-dicyclohexylcarbodiimide and its effect on URF13 sensitivity to fungal toxins. J Biol Chem. 268(8):5828-33.
[11] Korth KL, Levings CS 3rd (1993) Baculovirus expression of the maize mitochondrial protein URF13 confers insecticidal activity in cell cultures and larvae. Proc Natl Acad Sci U S A. 90(8):3388-92.
[12] Rhoads DM, Kaspi CI, Levings CS 3rd, Siedow JN (1994) N,N'-dicyclohexylcarbodiimide cross-linking suggests a central core of helices II in oligomers of URF13, the pore-forming T-toxin receptor of cms-T maize mitochondria. Proc Natl Acad Sci U S A. 91(17):8253-7.
[13] Siedow JN, Rhoads DM, Ward GC, Levings CS 3rd (1995) The relationship between the mitochondrial gene T-urf13 and fungal pathotoxin sensitivity in maize. Biochim Biophys Acta. 1271(1):235-40.
[14] Rhoads DM, Levings CS 3rd, Siedow JN (1995) URF13, a ligand-gated, pore-forming receptor for T-toxin in the inner membrane of cms-T mitochondria. J Bioenerg Biomembr. 27(4):437-45.
[15] Trotter, M et al. (2014) Cryptic genetic variation can make "Irreducible Complexity" a common mode of adaptation in sexual populations. Evolution 68, 3357-3367.
[16] Hecht, MH et al. (1990) De novo design, expression, and characterization of Felix: a four-helix bundle protein of native-like sequence. Science 249(4971): 8848-8891.
[17] Kamtekar, S et al. (1993) Protein design by binary patterning of polar and nonpolar amino acids. Science 262(5140): 1680-1685.
Autor: Andreas Beyer und Martin Neukamm


